|
Гипермаркет знаний>>Естествознание>>Естествознание 11 класс>> Межклеточный обмен веществами
5.5.6. Межклеточный обмен веществами
В настоящее время ещё не все детали поразительного по совершенству механизма воспроизводства генетической информации и молекулярного биологического синтеза установлены. Понято хотя бы главное, в том числе то, что каждая клетка в организме не создает всех нужных ей веществ, хотя генетической информацией обладает. Более эффективным оказывается путь специализации функций клеток, это приводит к совершенствованию организма в целом. Следовательно, между «элементалями» организма (клетками) должен непрерывно поддерживаться обмен, клетки и поставщики, и потребители потоков веществ. У простейших одноклеточных организмов процесс обмена веществами между клеткой и окружающей средой является процессом пищеварения, в результате которого происходит ассимиляция полезных веществ и удаление шлаков, остающихся после утилизации продуктов питания.
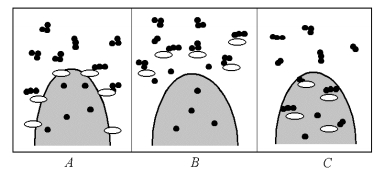
Рис. 5.26. Схема процессов ассимиляции:
А - мембранный; В - внеклеточный; С – внутриклеточный
За последние 10 - 15 лет удалось установить, что все разнообразие процессов ассимиляции полезных для клетки продуктов, свойственных живым организмам (от одноклеточных бактерий до млекопитающих), сводится к трем основным типам: внеклеточному, внутриклеточному и мембранному.
Схема соответствующих процессов приведена на рис. 5.26.
При внутриклеточной ассимиляции ферменты цитоплазмы гидролизуют (разлагают) небольшие молекулы питательных веществ (главным образом димеры - дисахариды, дипептиды и т.д.), проникшие в клетку путем диффузии или путем специального транспорта (эндоцитоза). Внеклеточное пищеварение осуществляется ферментами, которые выходят из цитоплазмы наружу. Здесь ферменты, как говорят биохимики, атакуют поглощенные организмом субстраты (крупные молекулярные комплексы), разрушают их до димерных или мономерных форм, которые уже способны затем проникать в клетку. У высших животных и человека этот процесс начинается в ротовой полости и продолжается в полости желудка, сочетаясь с внутриклеточным. Мембранная деполимеризация субстрата происходит на границе и имет черты как внеклеточного, так и внутриклеточного апроцессов.
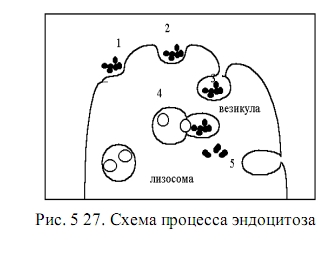
Оно малоэффективно в отношении крупных надмолекулярных комплексов. Поэтому для таких «лакомых кусочков» многие клетки используют цикл эндоцитоза.
Эндоцитоз начинается с того, что макромолекула питательного вещества прикрепляется к специальному мембранному белку, называемому рецептором. Рецепторы могут перемещаться по плазматической (полужидкой) мембране клетки, перенося с собой лиганды - прикрепленные к ним большие молекулы. Такой перенос оказывается более эффективным, чем случайная диффузия субстрата по мембране. За одну минуту циркулирующий белок может пройти расстояние около 10 мкм, что сравнимо с размерами некоторых клеток. Когда в одном месте собираются до тысячи рецепторов с лигандами, такой участок мембраны начинает впячиваться внутрь клетки и образует сначала ямку, а затем пузырек (везикулу).
Везикула отпочковывается от мембраны и перемещается в цитоплазму, где с ней соединяется лизосома с ферментами. Оболочка лизосомы предохраняет содержимое цитоплазмы от химического действия ферментов. Когда две оболочки входят в контакт и объединяются, ферменты переходят в везикулу и расщепляют пищевой субстрат, который поступает в цитоплазму, а белки оболочки везикулы возвращаются в мембрану клетки. Весь этот цикл происходит довольно быстро, за одну – две минуты.

Клетки различных организмов используют тот или другой тип ассимиляции, в зависимости от их эволюционного пути. Эндоцитоз и внутреннее пищеварение характерно для низших организмов, у высших животных и человека основные роли играют процессы полостного гидролиза и мембранного всасывания. Между ними существует функциональная связь, поскольку конечные продукты полостной деполимеризации служат исходным сырьем для мембранных процессов. В любом случае именно через мембрану клетки проходят встречные потоки веществ.
Оценка роли мембран возросла в последние годы, когда были начаты исследования механизма гормонального управления межклеточным обменом веществ. Например, известно, что клетки печени вырабатывают глюкозу в ответ на появление в крови гормона адреналина. Каким образом внешнее (по отношению к клетке) химическое воздействие запускает процесс выработки глюкозы?
Было предложено несколько моделей строения плазматической мембраны клеток, наиболее полной является твердокаркасная жидкокристаллическая модель. По ней основу всякой клеточной мембраны составляет двойной слой липидов (жирных кислот). Головные части молекул липидов гидрофильны, то есть хорошо смачиваются водой и водными растворами. Хвостовые цепи, наоборот - водоотталкивающие, гидрофобны, энергия их взаимодействия между собой превосходит энергию их взаимодействия с молеку-
лами воды. Головные части формируют внешние поверхности мембран, а гидрофобные концы обращены внутрь.
Бислой весьма стабилен и в то же время имеет свойства жидкости. В одну или сразу в обе поверхности бислоя встраиваются крупные молекулы белков различного назначения: циркулирующие (переносчики лигандов), коммутационные, рецепторы и эффекторы. С внутренней стороны мембраны имеется ажурный каркас с элементами дальнего порядка (свойственного кристаллам), он построен из микронитей (микрофиламент) спектрина, которые закреплены за некоторые коммутационные белки, встроенные в бислой мембраны.
Такая жидко-твердая структура допускает локальные и общие перестройки с различной степенью кооперативности. При этом структурное возмущение, распространяющееся по мембране после посадки гормона или медиатора (химического передатчика сигнала) на соответствующий только ему рецептор, приводит к изменению координации и каталитической активности других мембранно-связанных ферментов. По такому механизму передаются межрецепторные сигналы (взаимодействия). Например, инсулин через свой рецептор изменяет свойства совершенно другой белковой молекулы - 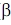 -адренорецептора. -адренорецептора.
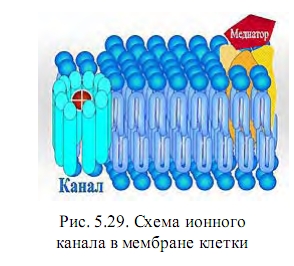
В мембране имеются каналы различного диаметра, стенками которых служат молекулы белков. Через эти каналы свободно могут проходить в обе стороны молекулы воды, некоторые ионы (натрия и калия) и некоторые кислотные остатки (фосфорных кислот, аминокислоты). Часть каналов открыта постоянно, другие открываются только тогда, когда связанные с ними ферменты расщепляют молекулы субстрата (при мембранном пищеварении).
В результате реакции конформация фермента заметно изменяется так, что канал открывается и отщепленные частицы могут войти в цитоплазму клетки (пример конформационного перехода). Сигнал на фермент натриевого канала может быть передан также от конформационного перехода на соседнем рецепторе.
Более сложные процессы передачи информации на уровне клетки происходят с участием мессенджеров (белков-посланников). В середине 80-х гг. выяснилось, что центральное место в передаче межклеточных управляющих сигналов занимают G-белки, такое название они получили из-за приспособленности для связи с гуаниновыми нуклеотидами (гуанин-пентоза 123фосфатными молекулами). G-белки прикреплены к внутреннему слою мембраны и могут передвигаться по нему от рецептора к эффектору.
Процесс начинается с того, что адреналин (или другой гормон) прикрепляется к наружнему концу своего рецептора. Выделяющаяся энергия связи передается на другом конце рецептора G-белку, это стимулирует обмен белка с присоединенным к нему гуаниндифосфатом. Обмен происходит с выделением энергии и G-белок диссоциирует, от него отделяется его компонент ?-субчастица (она несет ГТФ) и быстро мигрирует к белку - эффектору.
Это первый этап процесса.
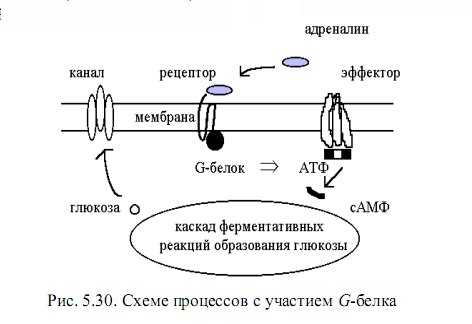
Второй этап начинается после того, как связывание -субчастицы с эффектором переводит в возбужденное состояние его электронную оболочку. В возбужденном состоянии химическая активность молекул повышается и становятся возможными новые реакции, в обычных условиях не протекающие. В рассматриваемом случае эффектор становится катализатором превращения аденозинтрифосфата (АТФ) в циклический аденозинметафосфат (сАМФ).
Активное состояние сохраняется до тех пор, пока в альфа-субчастице не произойдет гидролиз (отщепление одной фосфатной группы):
ГТФ > ГДФ (трифосфат > дифосфат)
Тогда -субчастица возвращается назад, возбужденное состояние эффектора снимается, и он прекращает создавать сАМФ. Таким образом, время «включенного» состояния эффектора задается временем протекания реакции гидролиза фосфатной цепи гуанинового нуклеотида в составе субчастицы G-белка. Можно говорить о том, что обнаружен механизм таймера или своеобразного реле времени на молекулярном клеточном уровне.
Наработанные за время активного состояния эффектора молекулы сАМФ запускают каскад ферментативных реакций образования глюкозы из животного крахмала - гликогена. Такие реакции давно и хорошо воспроизводятся в лабораторных условиях. Образованная глюкоза выходит через мембранные каналы в межклеточное пространство или в кровяное русло. Это ответ клетки на внешнее «послание» о необходимости для организма некоторого количества глюкозы.
Зачем природе потребовался такой (на первый взгляд усложненный) вариант передачи информации? Дело в том, что при таком механизме действия происходит большое усиление начального сигнала. Будучи в возбужденном состоянии эффектор преобразует сотни молекул АТФ на одну молекулу первичного, внешнего для клетки, мессенджера (в данном случае - адреналина), появляются сотни мессенджеров вторичных (до 500 в нашем случае) в цитоплазме клетки. За сопоставимое время через открытый канал мембраны успели бы пройти всего одна - две молекулы, и эффективность управления была бы низкой.
Процессы передачи межклеточного взаимодействия с участием G-белков имеют большое распространение в живых организмах, считают что они играют роль в различении запахов, когда количество воспринимаемых первичных ароматических молекул чрезвычайно мало. Эти белки могут регулировать работу ионных каналов в мембранах. Например, циклический сГМФ поддерживает в открытом состоянии натриевый канал. Gm-белок включает эффектор, который превращает сГМФ в нециклический ГМФ. После этого канал для ионов натрия закрывается и клетка в целом приобретает электрический заряд.
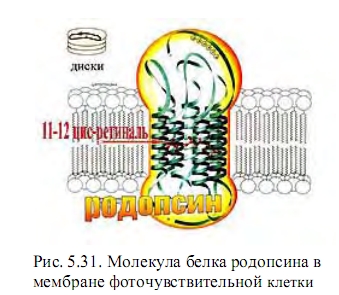
Мы можем теперь дополнить описание механизма зрения человека. В молекуле родопсина присутствует Gт-белок трансдуцин. Он контролирует белок-эффектор фосфордиэстеразу, которая превращает сГМФ в ГМФ. Процесс идет с усилением начального
сигнала (конформационного фотовозбуждения одной молекулы ретиналя) в 500 раз!
Кроме того G-белки служат своеобразными часовыми механизмами в организмах, время гидролиза ГТФ > ГДФ достигает нескольких секунд, то есть на уровне интервалов хорошо различаемых человеком.
Человек «чувствует» время потому, что в его клетках имеется такое важное соединение, как гуанинтрифосфат!
Концепции современного естествознания. Стародубцев В.А., 2-е изд., доп. — Томск.: Том. политех. ун-т, 2002. — 184 с.
Содержание урока
 конспект урока
опорный каркас
презентация урока
акселеративные методы
интерактивные технологии
Практика
задачи и упражнения
самопроверка
практикумы, тренинги, кейсы, квесты
домашние задания
дискуссионные вопросы
риторические вопросы от учеников
Иллюстрации
аудио-, видеоклипы и мультимедиа
фотографии, картинки
графики, таблицы, схемы
юмор, анекдоты, приколы, комиксы
притчи, поговорки, кроссворды, цитаты
Дополнения
рефераты
статьи
фишки для любознательных
шпаргалки
учебники основные и дополнительные
словарь терминов
прочие
Совершенствование учебников и уроков
исправление ошибок в учебнике
обновление фрагмента в учебнике
элементы новаторства на уроке
замена устаревших знаний новыми
Только для учителей
идеальные уроки
календарный план на год
методические рекомендации
программы
обсуждения
Интегрированные уроки конспект урока
опорный каркас
презентация урока
акселеративные методы
интерактивные технологии
Практика
задачи и упражнения
самопроверка
практикумы, тренинги, кейсы, квесты
домашние задания
дискуссионные вопросы
риторические вопросы от учеников
Иллюстрации
аудио-, видеоклипы и мультимедиа
фотографии, картинки
графики, таблицы, схемы
юмор, анекдоты, приколы, комиксы
притчи, поговорки, кроссворды, цитаты
Дополнения
рефераты
статьи
фишки для любознательных
шпаргалки
учебники основные и дополнительные
словарь терминов
прочие
Совершенствование учебников и уроков
исправление ошибок в учебнике
обновление фрагмента в учебнике
элементы новаторства на уроке
замена устаревших знаний новыми
Только для учителей
идеальные уроки
календарный план на год
методические рекомендации
программы
обсуждения
Интегрированные уроки
Если у вас есть исправления или предложения к данному уроку, напишите нам.
Если вы хотите увидеть другие корректировки и пожелания к урокам, смотрите здесь - Образовательный форум.
|
















